Aharoni, I. 1938. On some animals mentioned in the Bible. Osiris 5: 461-478.
Anderson, E. N. 2011. Ethnobiology: Overview of a growing field. In: Ethnobiology, edited by E. N. Anderson, et al.. Hoboken, NJ: Wiley-Blackwell.
Arment, C. 2022. Ruminating on created kinds and ark kinds. Answers Research Journal 15: 391-404. [PDF]
Atran, S. 1990. Cognitive Foundations of Natural History. Cambridge: Cambridge University Press.
Berlin, B. 1992. Ethnobiological Classification: Principles of Categorization of Plants and Animals in Traditional Societies. Princeton, NJ: Princeton University Press.
DeRemer, F. 2014. Structure, toledoths, and sources of Genesis. Journal of Creation 28(1): 53-58.
Gray, B. 2019. The #1 Mistake Most Everyone Makes Reading the Bible. 2nd edition. ebook. Spring Hill, TN: Walking the Text.
Hawley, L. 2015. The agenda of priestly taxonomy: The conceptualization of טמֵָא and שֶׁקֶץ in Leviticus 11. The Catholic Biblical Quarterly 77: 231-249.
Hunn, E. S. 2011. Ethnozoology. In: Ethnobiology, edited by E. N. Anderson, et al., 83-96. Hoboken, NJ: Wiley-Blackwell.
Hunn, E. S., and C. H. Brown. 2011. Linguistic ethnobiology. In: Ethnobiology, edited by E. N. Anderson, et al., 319-333. Hoboken, NJ: Wiley-Blackwell.
Kay, M. 2007. On literary theorists’ approach to Genesis 1: Part 2. Journal of Creation 21(3): 93-101.
McLain, M. A., M. Petrone, and M. Speights. 2018. Feathered dinosaurs reconsidered: New insights from baraminology and ethnotaxonomy. In: Proceedings of the Eighth International Conference on Creationism, edited by J. H. Whitmore, pp. 472-515. Pittsburgh, PA: Creation Science Fellowship.
Neville, R. 2011. Differentiation in Genesis 1: An exegetical creation ex nihilo. Journal of Biblical Literature 130(2): 209–226.
Ouro, R. 2002. Linguistic and thematic parallels between Genesis 1 and 3. Journal of the Adventist Theological Society 13(1): 44-54.
Ross, N., and C. Revilla-Minaya. 2011. Cognitive studies in ethnobiology: What can we learn about the mind as well as human environmental interaction? In: Ethnobiology, edited by E. N. Anderson, et al., 335-349. Hoboken, NJ: Wiley-Blackwell.
Sarfati, J. D. 2015. The Genesis Account. Powder Springs, GA: Creation Book Publishers.
Sanders, R. W., and K. P. Wise. 2003. The cognitum: A perception-dependent concept needed in baraminology. In: Proceedings of the Fifth International Conference on Creationism, edited by Robert L. Ivey, 445-455. Pittsburgh, PA: Creation Science Fellowship.
Simkins, R. 1991. Yahweh’s Activity in History and Nature in the Book of Joel. Ancient Near Eastern Texts and Studies, Vol. 10. Lewiston, NY: The Edwin Mellen Press.
Strawn, B. A. 2009. kĕpîr ʾărāyôt in Judges 14:5. Vetus Testamentum 59: 150-158.
Wenham, G. J. 1987. Word Biblical Commentary. Vol. 1. Genesis 1-15. Nashville, TN: Thomas Nelson.
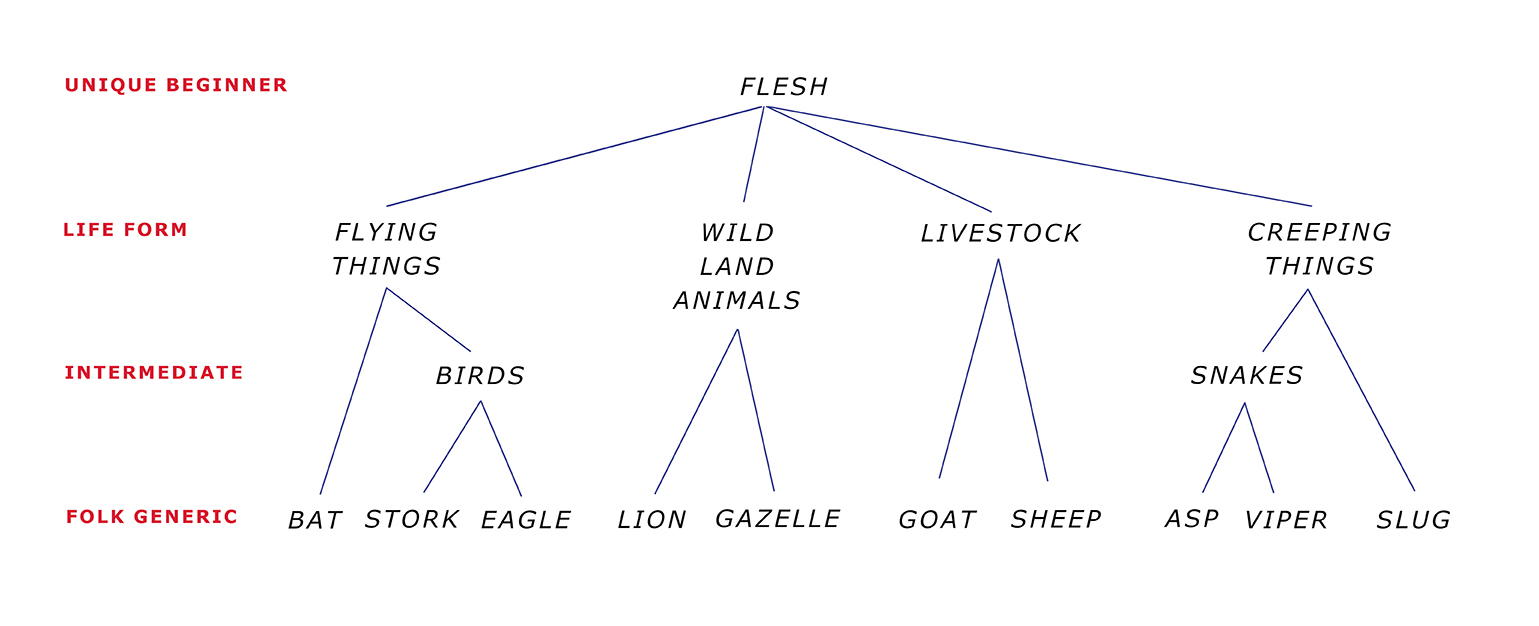
Whitekettle, R. 2001. Where the wild things are: Primary level taxa in Israelite zoological thought. Journal for the Study of the Old Testament 93: 17-37.
Whitekettle, R. 2002. All creatures great and small: Intermediate level taxa in Israelite zoological thought. Scandinavian Journal of the Old Testament 16(2): 163-183.
Whitekettle, R. 2003. Of mice and wren: Terminal level taxa in Israelite zoological thought. Scandinavian Journal of the Old Testament 17(2): 163-182.
Whitekettle, R. 2008. Forensic zoology: Animal taxonomy and rhetorical persuasion in Psalm l. Vetus Testamentum 58: 404-419.
Wood, T. C., K. P. Wise, R. Sanders, and N. Doran. 2003. A refined baramin concept. Occasional Papers of the Baraminology Study Group 3: 1–14.