From Polar Bears to Pandas
Post-Flood Diversification of the Bear Kind
Chad Arment (2021)

American Black Bear, Ursus americanus (Yellowstone NPS / Neal Herbert)
There were no polar bears on the Ark. There weren’t any giant pandas, either. In fact, all modern bear species are descended from a very different ancestral pair that was on the Ark. In this article, I’ll give a general overview of the bear kind after the Flood, and how different species developed as they spread around the world. We’ll also take a closer look at some of the more interesting specializations that occurred as species adapted to post-Flood environments.
Today, bears can be found in Europe, Asia, North America, and South America. Modern species include three subfamilies within the Family Ursidae. There are additional subfamilies that are now extinct. There are eight living species of bears: Asiatic black bear (Ursus thibetanus), American black bear (Ursus americanus), sun bear (Helarctos malayanus), sloth bear (Melursus ursinus), brown bear (Ursus arctos), polar bear (Ursus maritimus), Andean bear (Tremarctos ornatus), and giant panda (Ailuropoda melanoleuca).

Polar Bear, Ursus maritimus (USFWS / Terry Debruyne)
Baraminic Boundaries
Do bears make up their own created kind? In other words, were the ancestors of modern bears that appeared during the creation week ‘true bears,’ and only ancestral to bears? Or were they ancestral to a wider range of creatures? Or are there more than one created kind of bears? The creationist studies that have included bears show that the Ursidae are monobaraminic (meaning they share enough similarities to likely have a common ancestor), but it is not always clear whether they are apobaraminic (significantly discontinuous from other creatures) (Hennigan 2010; Wood 2021). The Ursidae would need to be both monobaraminic and apobaraminic to be characterized as holobaraminic. The morphological traits that delineate the Ursidae (their synapomorphies, primarily in the basicranium) can be found in Wozencraft (1989).
One of the best ways to determine that living bears, at least, are monobaraminic is through hybridization studies. A few hybrids have been found in the wild, but because bears are popular animals for zoos and other animal exhibits, there have been a number of cases of accidental hybridization. Known hybrids include Ursus arctos x Ursus maritimus (Preuß et al. 2009; Pongracz et al. 2017), Ursus americanus x Ursus arctos (Scherren 1907; Van Gelder 1977), Ursus arctos x Ursus thibetanus (Van Gelder 1977), Tremarctos ornatus x Ursus thibetanus (Mondolfi and Boede 1981), and Helarctos malayanus x Melursus ursinus (Asakura 1969). A likely Helarctos malayanus x Ursus thibetanus wild hybrid has also been noted (Galbreath et al. 2008). Secular attempts to create an evolutionary phylogeny of bears have run into problems in the past because they didn’t adequately take into account gene flow between the different species, especially in the nature of transposable elements (Kumar et al 2017; Lammers et al 2017). The giant panda is the only living bear species that hasn’t hybridized with other bears. Statistical baraminology, however, based on morphological traits, along with fossil evidence, supports the giant panda’s placement within the bear kind.
Evolutionists believe that bears evolved from basal arctoids (Infraorder Arctoidea), from which also evolved mustelids (weasels), mephitids (skunks), procyonids (raccoons), ailurids (lesser pandas), and pinnipeds (seals). A creationist researcher could theorize that all arctoids are one kind, or that each family (more or less) is a distinct and separate kind. It doesn’t really matter which theory is correct. Each incorporates significant change over the same time period within the kind (the holobaramin). For now, we will focus on bears (Family Ursidae) as a monobaramin. With that premise, we know that one pair of animals on the Ark was ancestral to all living bears.

Giant Panda, Ailuropoda melanoleuca (© Cedar)
Post-Flood Diversification
The fossil record of bears is consistent with a post-Flood record. As with most modern mammals, there do not appear to be any pre-Flood bears in the Flood rocks, just as there are no human fossils. After the Flood, from a single pair of animals on the Ark, its offspring spread around the world and diversified, splitting into numerous lineages. There are four major lineages within the Ursidae: the Ursavinae (the extinct 'first bears'), the Ailuropodinae (giant pandas), the Tremarctinae (short-faced bears), and the Ursinae (typical bears). Additional extinct lineages are sometimes included within the Ursidae: the Amphicynodontinae and the Hemicyoninae (dog-bears).
I’ll note here that throughout this article I use the terminology for a secular timeframe (Pleistocene, Pliocene, etc.). While the evolutionary model uses these within a framework involving millions of years, within a creation model these simply refer to chronological periods after the Flood. (We can consider them 'snapshots' of migration and adaptation after the Flood, fulfilling God's intent for creatures to fill the earth again.) We see, in the post-Flood fossil record, very distinctive developmental trajectories up to modern times. This timeframe for ursids is after the Flood, but no argument is being made here for a specific post-Flood boundary.

Brown Bear, Ursus arctos (© Slowmotiongli)
Amphicynodontinae (Early Arctoids)
There is no consensus in evolutionary biology regarding this (possibly paraphyletic) group’s relationship to bears. Qiu (2003), for example, treats them as a ‘separate family of the arctoid carnivorans, comprising four subfamilies: Daphoeninae, Temnocyoninae, Haplocyoninae, and Amphicyoninae.’ Consequently, while a creationist could consider these early members of the bear kind, they may comprise a different kind altogether (or more than one kind). In the fossil record they are found in Late Eocene layers of North America (Bryant 1993; Van Valkenburgh 1994), throughout the Oligocene of North America, Europe, and Asia (Val Valkenburgh 1994; Morlo and Nagel 2007; Gardin et al. 2021), and into the Miocene of North America (Hunt, Jr. 1998). These were generally raccoon- or otter-sized.
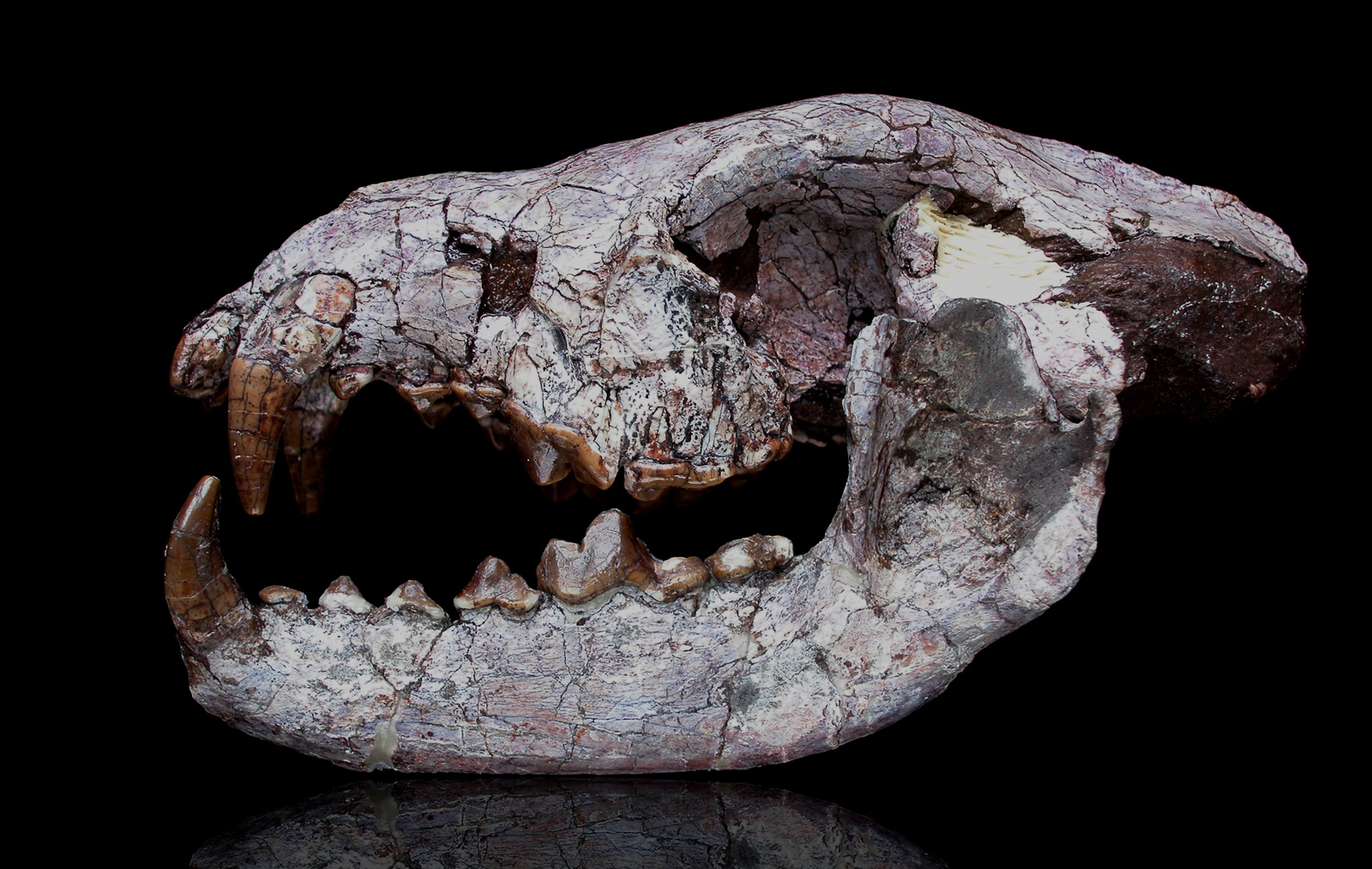
Research is spotty on these creatures’ biology. Gardin et al. (2021) noted that the genet-sized Amphicynodon leptorhynchus of Europe had forelimb anatomy indicating strong climbing and grasping abilities, so it may have been arboreal. The North American otter-like Kolponomos appears to have had a littoral lifestyle, along the Pacific Coast, using its incisors and canines to pry and perhaps crush mollusk shells (Tseng, Grohé, and Flynn 2016).

Kolponomos skull (USNM PAL215070 / SI.edu)
Hemicyoninae (Dog-Bears)
Some dog-bears, like the Oligocene Cephalogale, were raccoon-sized, while others were as large as our modern bears (McLellan and Reiner 1994). Some species may have occupied “ecological niches close to those of some extant Canidae. . . . The jaws display a balance between shearing and chopping teeth like in canids . . .” (De Bonis 2012). Some species were well adapted for cursorial predation. Their eventual extinction may have been due to competition with other bears, or even with large felid species.
This extinct group is sometimes placed in its own family. From a creationist perspective, it really doesn’t matter if it is part of the bear kind or is just another kind in the creationist ‘orchard.’ (Note: these ‘dog-bears’ are not the same creatures as the also-extinct ‘bear dogs’ of the Amphicyonidae, which in turn are no relation to the previously noted Amphicynodontinae.) Hemicyonines first show up in Oligocene layers (Europe and Asia), with numerous species throughout Miocene layers (Europe, Asia, North America). There are some early papers referring to Pliocene fossils, but those appear to belong to species later moved to different subfamilies. There are reported Cephalogale from the Chinese Eocene, but these are unconfirmed as the specimens are missing (Qiu 2003). There are three ‘tribes’ of genera (approx. 32 species) that diversified during their duration of post-Flood life.
It has been suggested that Cephalogale is basal to both other Hemicyoninae and to the Ursavini, which are considered the earliest true bears (McLellan and Reiner 1994; Wang, McKenna, and Dashzeveg 2005; Sotnikova et al. 2021).
Miocene hemicyonine Phobergale shareri, holotype (Xiaoming Wang, CC-BY 3.0)
Ursavinae
The Ursavinae (which we can narrow down to the tribe Ursavini) consists of three genera: Ballusia, Ursavus, and the recently described Aurorarctos. These are small bears, smaller species close to wolverine-sized, larger species being about wolf-sized (De Torres Perezhidalgo 1992; Qiu et al. 2014). These are the earliest ‘true bears,’ Ballusia and Ursavus showing up in Early Miocene deposits in Europe, later in Asia (Sotnikova et al. 2021). Ursavus is also found in later Miocene deposits in North America. Qiu et al. (2014) offered cladistic support that Ursavus (sometimes referred to as the ‘dawn bear’) was closer to modern bears than Ballusia. The North American Aurorarctos, so far only known from late Middle Miocene Nebraska (Jiangzuo and Flynn 2020b), was an arboreally adapted herbivore-omnivore, which is interesting as the smaller living bears are usually good tree-climbers.
From a creationist standpoint, the Ursavini may be very close morphologically to the Ark kind pair, or may have descended from the Hemicyoninae. Because most North American records are later than Eurasian fossils, it seems likely the North American populations were due to early post-Flood migration. However, a questionable fossil from the John Day Formation in Oregon (Qiu et al. 2014) might indicate Ursavus developed first in North America (possibly from hemicyonine stock?) and then migrated to Eurasia. (Old World-New World migrations would have occurred via the Bering Land Bridge.)
In any case, Ursavus was a key genus (or a paraphyletic group of related genera) in the development of modern bears, as it is likely modern species have their origin through Ursavus species in Asia (Wang et al. 2017). (More on that below.)

Ursavus tedforti skull cast (Skye M., CC-BY-SA 4.0)
Ursinae (Typical Bears)
Traditionally, Ursus minimus was considered to be the earliest Ursus species to develop from Ursavus, first appearing in Europe in the Pliocene (McLellan and Reiner 1994). It was very similar to the modern Asian black bear in appearance. For some time, the taxonomy of early fossil bears was argued by lumpers who thought U. minimus included a number of subspecies, while others split it into several species (Baryshnikov and Lavrov 2013). In addition, some researchers suggested that ‘black bears’ (extant and fossil) should be placed in either of two separate genera: Euarctos and Protarctos (Wang et al. 2017), with Euarctos minimus being just one of several early bears, and Euarctos including the living Asian and American black bears (Qiu, Deng, and Wang 2009). Baryshnikov and Lavrov (2013) note that the distinction between Euarctos and Ursus is primarily over the morphology of a tooth, so Ursus is still preferred by many; I’ll continue to use Ursus here.
Protarctos abstrusus is a Pliocene North American ursine (known from Idaho and the Canadian Arctic) closely related to Ursavus tedfordi, the most ‘advanced’ of that stem genus (Wang et al. 2017). While P. abstrusus had been considered a possible direct ancestor to the American black bear (McLellan and Reiner 1994; Krause et al. 2008), Ursus americanus is now believed to have been part of a more recent (Early Pleistocene) migration to North America (Wang et al. 2017). P. abstrusus was a basal ursine, and Protarctos as a genus is considered ancestral to modern bears.
The Ursinae includes several distinct lineages of bears which we will consider: the black bears, the sloth bear, the cave bears, and the brown bears (which includes the polar bear).

American Black Bear cub, Ursus americanus (© Tony Campbell)
The Black Bears

Asian Black Bear, Ursus thibetanus (© Vova Pomortzeff)
We know that the extant Asian black bear (Ursus thibetanus) is found in Pliocene and Pleistocene fossil sites in Eurasia, and that it has a very strong resemblance (though not exact) to Ursus (Euarctos) minimus (Baryshnikov and Zakharov 2013), suggesting it is a close descendant. Future fossil discoveries may clarify that point. It seems likely that hybridization was not uncommon between the two species, particularly early on. The American black bear (Ursus americanus) developed from Asian black bears that migrated to North America in the Early Pleistocene (Puckett et al. 2015; Wang et al. 2017).
The sun bear (Helarctos malayanus) lives in the tropical and subtropical forests of south and southeast Asia, though had a wider range in Asia historically (Meijaard 2004; Li et al. 2017). Its fossils first show up in the Asian Pleistocene (Meijaard 2004), and suggest a distinct emergence from earlier U. minimus-U. thibetanus populations.

American Black Bear, Ursus americanus, with a cinnamon pelage (© Rimal5)

Sun Bear, Helarctos malayanus (Robert Elsmore)
The sloth bear (Melursus ursinus) is a highly adapted bear that specializes in a social insect (ant and termite) diet (as much as 50% (McNab 1992)), though also eats sugar-rich fruits (Seidensticker, Yoganand, and Johnsingh 2011). Sloth bear remains have been found in Pleistocene caves in India (Prasad 1996). Most phylogenetic analyses place sloth bears as sister taxa to both the black bear-sun bear and brown bear-polar bear lineages, so it is likely that they developed as a distinct lineage from U. minimus-type ancestry. Diet may have driven the specialized adaptation that led to the emergence of Melursus as a distinct lineage; other animals on the Indian subcontinent likely filled predatory and tuber/root harvesting niches that other bears have taken (Laurie and Seidensticker 1977).

Sloth Bear, Melursus ursinus (© Vladimir Ivanov)
The Cave Bears
The best-known cave bear, Ursus spelaeus, is just one of several species in the cave bear lineage, found in Pleistocene deposits from western Europe to northeast Siberia (Knapp 2019). The exact developmental route is still debatable, but cave bears appear to have arisen from Ursus etruscus (Hänni et al. 1994), which likely emerged from Ursus minimus. The earliest species with cave bear traits was Ursus dolinensis of Early Pleistocene Europe (García and Arsuaga 2001), with cave bears transitioning through the Middle Pleistocene form Ursus deningeri into the spelaus lineage of the Late Pleistocene. (Related cave bears include U. ingressus, U. kudarensis, and the ‘small cave bears,’ among others; there is some debate over where these branch off, or even if they should be considered distinct species (Stiller et al. 2014).)
Cave bears seem to have started out small (as their ancestral U. deningeri was), with some branches only diverging into extremely large bears later (Hofreiter et al. 2002). While some may have been herbivorous, others appear to have adapted to omnivorous or carnivorous lifestyles (Knapp et al. 2009). In one locality, two distinct cave bears (U. ingressus and U. spelaeus eremus), along with the brown bear (U. arctos) lived side-by-side, suggesting niche partitioning between the species (Bocherens et al. 2011).
What is most interesting about cave bears is that they died out by the end of the Pleistocene, while their contemporaries, the brown bears, continued to thrive. Climate change, habitat fragmentation, predation by other carnivores, and human hunting may all have put pressure on dwindling populations (Stiller et al. 2014; Terlato et al. 2019). Ancient DNA demonstrated a population decline before its extinction; increasing use of caves by humans may have instigated deadly conflicts for the bears (Gretzinger et al. 2019). Some cave bear DNA still survives, though, in brown bears, showing that the species hybridized in the Pleistocene (Barlow et al. 2018).

Cave Bear, Ursus spelaeus, skull (James St. John, CC-BY)
The Brown Bear
Brown bears (Ursus arctos) are a sister clade of the cave bears (Barlow et al. 2018). From the early Ursus minimus-like bears arose the Etruscan bear, Ursus etruscus, which populated Europe, Asia, and North Africa from the Pliocene to the Pleistocene. It appears to have given rise, separately, to both the cave bear and the brown bear lineages in the Pleistocene (Waits et al. 1998). Brown bears have an extensive modern Holarctic distribution in Europe, Asia, and North America, which was historically even greater. The brown bear was formerly found in northern Africa (though whether it should be referred to as a distinct subspecies, the Atlas bear, is debatable (Calvignac et al. 2008)), while the grizzly was found as far east in North America as Ohio, Kentucky, and Ontario (Guilday 1968) and into the Southwest and northern Mexico (White 1965; Trevino and Jonkel 1986; Gallo-Reynoso 2008; Czaplewski and Willsey 2013).
Genetic research shows that early European brown bears were more flexible in hibernation behavior than their close kin, the cave bears (Fortes et al. 2016). (The latter demonstrated high fidelity to their birth caves.) Their ability to adapt likely played a role in their survival to modern times. Numerous genetic lineages have been mapped with brown bears, testifying to their adaptability. One such lineage led to the emergence of the polar bear (Shields et al. 2000).

Brown Bear, Ursus arctos (© Anita Huszti)
The Polar Bear
Polar bears have a circumpolar distribution, primarily found in the Arctic regions of North America, Europe, and Asia. They express a number of adaptations that set them apart from other bears. First, they are hypercarnivores with specialized craniodental characteristics, feeding mainly on seals and especially on seal blubber, being physiologically adapted to a lipid-rich diet (Rinker et al. 2019; Castruita, Westbury, and Lorenzen 2020). Their claws are adapted to facilitate moving on ice and securing prey, they have smaller tails and ears to reduce heat loss, they acquire substantial adipose deposits under their skin for thermal regulation and energy reserves, and they have enlarged front paws and denser limb bones to aid swimming (Slater et al. 2010; Liu et al. 2014; Rinker et al. 2019). The most obvious adaptation, of course, is the lack of pigment in their fur, providing camouflage in a snowbound environment.
Polar bears (Ursus maritimus) diverged from within a brown bear (Ursus arctos) lineage (Shields et al. 2000; Lindqvist et al. 2010; Nakagome, Mano, and Hasegawa 2013). The secular model is still debating when this occurred (due to too few polar bear fossils), but generally accepts that it was a ‘recent’ divergence (to them, meaning less than 500,000 years (Castruita, Westbury, and Lorenzen 2020)) resulting in rapid morphological adaptation to an Arctic environment. The oldest known polar bear fossil (subfossil, technically) is dated within a secular timeframe to 110,000 to 130,000 years old (Ingólfsson and Wiig 2008). For a creationist, this is also a recent development, occurring within the Ice Age.
Polar bears have low genetic diversity compared to brown bears, mostly due to a smaller effective population size (Castruita et al. 2020). Even after species diverged, however, there is evidence of significant gene flow from introgressive hybridization, from polar bear to brown bear, especially in North American populations (Cahill et al. 2013; Liu et al. 2014). The brown bear population in Alaska’s Admiralty, Baranof and Chichagof Islands still shows the genetic markers of polar bears present there when they colonized the islands (Cahill et al. 2015). Interestingly, this does suggest that recent media attention on ‘the plight of the polar bear’ due to hybridization with brown bears (resulting in ‘grolar bears’ or ‘pizzlies’) is little reason to worry about brown bear genes swamping polar bear populations. The gene flow is into the brown bear population, rather than into the polar bears. There are a few possible explanations for asymmetric gene flow (Cahill et al. 2015), but certainly there are environmental pressures that would result in ‘brown bear’ traits being too detrimental for survival in an Arctic habitat. Some researchers do suggest that there were introgressive hybridization events from both brown and black bears into the ancestors of modern polar bears, which may have provided useful novel traits during times of climatic change (Lan et al. 2016).
Researchers have noted that selective pressures on variation already present in ancestral alleles and for new mutations appear to have occurred within the polar bear genome (Castruita et al. 2020). While some of these adaptations are explainable through processes like natural selection and genetic drift, some of these adaptations are intriguingly complex. Polar bear hair is particularly interesting. It doesn’t simply lack pigment. The guard hairs are porous and hydrophobic, providing excellent thermal insulation both in air and underwater (Shao, Wang, and Bai 2020). The fractal porosity of the hair’s inner structure has led some to suggest the polar bear ‘evolved in a perfect mathematical way,’ to explain its superior thermal protection (He, Li, and Wang 2016). A better explanation would be that the bear kind was created to rapidly adapt to new environments. Early attempts to show that polar bear hair was capable of transmitting UV light to the bear’s skin in a fiber-optic manner were critiqued by a number of scientists (e.g. Koon 1998), and the idea was dropped. A more recent paper, however, suggests that while a single hair may not work effectively, taken collectively the hairs’ optical properties are worth further examination (Khattab and Tributsch 2015).

Polar Bears, Ursus maritimus, feeding on dead whale (© Gail Johnson)
Tremarctinae (Short-Faced Bears)
The majority of genetic and morphological studies indicate that the tremarctines are a sister clade to ursines (Kutschera et al. 2004; Krause, et al. 2008; Kumar et al. 2017), probably splitting off prior to the major ursine radiation. [There are some who suggest that the Tremarctinae should be with the Ursinae (Qiu et al. 2014), but this argument isn’t well developed.] There isn’t enough fossil material to determine the exact route, but Ursavus is assumed to be a probable ancestor (McLellan and Reiner 1994). All tremarctine fossils are from the New World, probably first diverging in North America. The earliest tremarctine is Plionarctos, with records in the late Miocene of Oregon and the Late Miocene to Early Pliocene Gray Fossil Site in Tennessee (Tedford and Martin 2001; Wallace and Wang 2004). That subsequent genera (Arctodus, Arctotherium, and Tremarctos) arose from Plionarctos itself, appears to be a reasonable supposition (Tedford and Martin 2001).
The only living tremarctine bear is the Andean (or spectacled) bear, Tremarctos ornatus, which is only found in the Andes of South America. Tremarctos originated in North America in the Pliocene, likely from Plionarctos. Arctodus also appears to have diverged, separately, from Plionarctos in the Late Pliocene (Soibelzon, Tonni, and Bond 2005), with fossils found throughout North America into the Pleistocene (Soibelzon et al. 2008). The giant South American short-faced bear Arctotherium either diverged from Tremarctos, or had a common ancestor with Tremarctos that didn’t give rise to Arctodus (Schubert et al. 2019). Arctotherium first shows up in El Salvador in the Pliocene, likely as part of the ‘Great American Biotic Interchange’ (Soibelzon et al. 2008). This suggests that its divergence was in North America, and the genus eventually made its way to expand throughout eastern South America. Arctotherium fossils in South America are exclusively Pleistocene (Soibelzon, Tonni, and Bond 2005).
The scenarios given in that last paragraph may be revised with new research (Salis et al. 2021). While neither Arctodus and Arctotherium appears to have hybridized with their contemporary Pleistocene ursine neighbors, the North American Tremarctos apparently did back in the Pleistocene. There is also evidence that Tremarctos hybridized with Arctotherium in the Late Pleistocene, after the ancestors of Arctotherium dispersed towards South America. The authors suggest that the hybridization data suggests Tremarctos is genetically closer to Arctodus; additional research may clarify the issue.
Arctodus simus in North America and Arctotherium angustidens in South America were both extremely large short-faced bears, though not closely related. Both may have weighed over a metric ton (Schubert et al. 2010; Soibelzon and Schubert 2011). Their giant morphologies were the result of convergence, not relationship (Mitchell et al. 2016). Both appear to have been omnivorous (Figueirido and Soibelzon 2009; Donohue et al. 2013), rather than hyper-scavengers or cursorial predators as some early researchers suggested. Their large sizes may have contributed to their demise, as smaller species with ‘conservative morphologies’ managed to survive the Pleistocene (Soibelzon and Schubert 2011).
Andean bears occupy a wide range of habitats across the Andes. They are medium-sized bears, and omnivores that eat a lot of plant material and fruits (though the oldest fossil Andean bear found in a Peruvian cave may have been more carnivorous (Stucchi et al. 2009)). Like all short-faced bears, they have a massive, blunt skull and exhibit short muzzles (García-Rangel 2012). Many show the ‘spectacles,’ or white to light yellow masking on their face and around their eyes. One interesting aspect of their morphology is the presence of a false thumb, though more constricted than is found in giant pandas. It doesn’t seem to be used in feeding, but may help in climbing (Salesa et al. 2006a). Its presence does not indicate a close relationship to giant pandas, but rather may point to it as a character found in ancestral species that was lost in the ursine lineages and adapted to a new use in giant pandas.

Andean Bear, Tremarctos ornatus (© Thorsten Spoerlein)
Ailuropodinae (Giant Pandas and Kin)
For years there was debate over whether the living giant panda (Ailuropoda melanoleuca) is a bear or if it is related to the lesser pandas (Family Ailuridae). After all, they are bamboo-specialists, have morphological traits that seem more ‘lesser panda’-like (particularly the ‘false thumb,’ an enlarged radial sesamoid bone), and behave very differently than typical bears (O’Brien et al. 1985; Vallittu et al. 2021). While morphological evidence for a bear identity was strongly demonstrated previously (Mayr 1986), it took molecular and cytological research in the 1980s to confirm for most researchers that the giant panda was a bear, though one demonstrating remarkable divergence and rapid adaptation within the Ursidae (O’Brien et al. 1985).
There are two major branches within the ailuropodine lineage (Qiu et al. 2014): 1) Kretzoiarctos, which likely developed into Indarctos and Agriotherium, and 2) the ‘giant panda’ lineage: Agriarctos, Ailurarctos, and Ailuropoda. The European Middle Miocene Kretzoiarctos precedes all other known ailuropodines (Abella et al. 2012), but a scanty fossil record muddles its position in regard to the giant panda lineage. Further fossil discoveries will hopefully clarify details.
Indarctos shows up first in Miocene Europe. It migrated to Northern Africa, Asia, and North and Central America (Qui 2003; Abella et al. 2012). Viranta (2004) notes that Indarctos “were the first large-bodied bears with omnivorous tendencies,” as previous bears had dentition best suited for carnivory. Omnivory may have been an adaptation allowing the bear to move between a plant dominated diet and meat dominated diet as the seasons changed. Indarctos had only a very small ‘false thumb,’ which is missing entirely in its descendant, Agriotherium (Salesa et al. 2006b). While there are a number of Miocene fossils, Indarctos doesn’t appear to have survived into the Pliocene (Baryshnikov and Tleuberdina 2017; Abella et al. 2019; Jiangzuo and Hulbert Jr. 2021). (The ‘Pliocene’ Kimball Formation given in Schultz and Martin (1975), should be considered Miocene Ash Hollow Formation (Diffendal 1990).)
Miomaci was a smaller offshoot of Indarctos that shows dental microwear similar to that of the bamboo-eating giant panda (De Bonis et al. 2017). Miomaci was not directly related to the giant pandas, but its herbivory suggests that the ailuropodine lineage was adaptationally flexible, allowing for certain unique lifestyles to emerge more than once.
There were at least nine species of the extinct giant short-faced bear Agriotherium. This genus had, in the past, been placed in other subfamilies, such as the Hemicyoninae (Qiu 2003), or in its own subfamily, the Agriotheriinae. Currently, researchers place it as an early offshoot from Indarctos (Miller and Carranza-Castañeda 1996; Jiangzuo and Flynn 2020a) in the Old World, or at least closely related (Qiu et al. 2014), with both genera subsequently migrating to the New World separately. With long legs, and a light build, these short-faced bears were among the largest carnivorous mammals known. There is some debate over Agriotherium’s diet: Sorkin (2006) suggested they most likely included a lot of plant material in their diet and may have roamed large home ranges looking for animal carcasses to scavenge. One smaller species recently described from North America has dentition that suggests a more carnivorous diet (Jiangzuo and Flynn 2020a). A dental microwear study on the African Agriotherium suggests it focused on soft tissue and bone, but was unable to say if this was from predation or scavenging (Stynder et al. 2019).
Agrotherium fossils are found in Miocene Asia (Qiu 2003; Ogino et al. 2011), and Pliocene Europe, Asia, and North America (Qiu 2003; Martin 2013). In North America, fossil have been found ‘from California to Florida and from Nebraska to southern Mexico’ (McLellan and Reiner 1994); the earliest being from the Bone Valley Formation near Brewster, Florida, (Everhart and Hawkins 2020) which is considered Middle Miocene to Early Pliocene. The latest uncontested North American fossil may be from the Blancan (Late Pliocene) (Samuels et al. 2009). Agriotherium africanum is found in Late Miocene/Early Pliocene layers of South Africa (Hendey 1977; Sorkin 2006). While some fossils are found in the latest Pliocene sediments, I’ve not yet located a categorically Pleistocene record for Agriotherium (though some publications hint at such).
The fossil evidence suggests that Ailuropoda (the giant panda) derives from the Asian Late Miocene Ailuractos, which in turn has many similarities to, and probably emerged from, Ursavus (Jin et al. 2007; Figueirido et al. 2011). The Late Miocene Agriarctos is part of this ‘giant panda’ lineage, but there is a lack of suitable fossil material to make any conclusions about its placement (Abella et al. 2012). It may simply be an early offshoot. The earliest ‘true’ giant panda is known from the late Pliocene, Ailuropoda microta (Dong 2008), and is smaller than the modern species.

Giant Panda, Ailuropoda melanoleuca (© Cedar)
Are all bears really post-flood?
In Arment (2020b), I showed that (among other arguments) the sheer number of carnivore genera crossing the Pliocene-Pleistocene boundary around the world essentially nullifies that location as a reasonable Flood/post-Flood boundary. Seven genera within the Ursidae alone are found on both sides of that boundary. Additional arguments in the paper point out that terrestrial Miocene fossils (where the bulk of ursid diversification occurs) should be considered post-Flood (see also Arment 2020a). As the only serious effort to push an upper Cenozoic post-Flood boundary is clearly flawed, creationists should focus their Flood boundary efforts elsewhere. (I’m not making any arguments regarding non-fossiliferous sediments or primarily marine sediments.) A lower (perhaps non-contiguous) boundary makes more sense of the fossil evidence.
If the Family Ursidae is a monobaramin, then its fossils are found in post-Flood sediments. Hybridization within the kind (and including with extinct bears, as noted with cave bears and short-faced bears), suggests that bears are a monobaramin. So there was only one pair of ursids (or ursid ancestors) on the Ark, and modern bears are the result of post-Flood diversification.
One attempt to salvage an upper boundary is the idea that what appear to be the same species or genera above and below the proposed boundary are actually not: that characteristics might appear in a modern population that originally appeared in a fossil population, even if they didn’t occur in the pair of animals saved on the Ark. The problem with this is that species and genera aren’t diagnosed with generic characters. Diagnostic character traits involve suites of characters and synapomorphies that are found only in those developmental pathways. They don’t just appear and reappear out of the blue. The development of modern ursid genera and species can be traced very effectively in the fossil record, primarily through skull and dental characteristics.
a creationist summary of bear diversification
Focusing on the ‘true bears’ here, based on their fossil record, bears diverged most significantly soon after leaving the Ark, continued to diversify as they spread to different continents and moved into empty niches, and the species we know today began emerging from those lineages during the Pleistocene. We can summarize these events as follows:
- Miocene: The Ursavini appear in Early Miocene deposits, with relevant genera Ballusia and Ursavus emerging. Kretzoiarctos appears in late Middle Miocene, along with related Indarctos, while Agriotherium, Ailurarctos, and Agriarctos appear Late Miocene. Plionarctos (the first short-faced bear) develops in Late Miocene North America, likely from Ursavus.
- Pliocene: Protarctos and Ursus emerge in the Pliocene from Ursavus. Short-faced bear genera Tremarctos, Arctodus, and Arctotherium diverge from Plionarctos. Ailuropoda appears in Asia.
- Pleistocene: Within the Pleistocene, American black bears, sun bears, sloth bears, cave bears, brown bears, and polar bears appear.
Within a secular timeframe the Pleistocene marks the start of several Ice Ages. For creationists, there is only one Ice Age, and the warm period that follows the Flood is gradually building up to it. Essentially, the energetic Flood produced warm oceans that began cooling, and over time ice began to build up at the poles, eventually spreading to the peak of the Ice Age (glacial maximum), at which point the global climate gradually warms up to modern temperatures.
Climate was incredibly important to the diversification of bears. (Even secularists recognize the importance of environmental changes at the Miocene-Pliocene boundary for bear diversification (e.g. Krause et al. 2008).) The earliest diversification introduced the greatest amount of change and produced several major lineages. Bears were able to cross into different continents, including North America via the Bering Strait, and target new ecological niches. This led to speciation, but also to hybridization between different lineages. This allowed beneficial newly emerged traits to move to different populations. The early centuries after the Flood show the greatest diversification within the kind.
Competition with other species (particularly other predators) and other selective pressures influenced the emergence of diverse phenotypes, some of which survived and others which disappeared. The development of new behaviors like denning also drove divergence. As Fowler et al. (2021) note, “Behavioural plasticity in response to environmental change contributes to diversification by exposing individuals to novel conditions through their interactions with resources or dispersal to new areas.” During the Pliocene, the Isthmus of Panama arose as a land bridge to South America, allowing some bears to move in that direction. The Bering Strait land bridge to North America remained open until the route was blocked by the ice sheets late in the Ice Age. (Melting glaciers allowed the route to open briefly again, but rising sea levels eventually flooded the land bridge for good.) The speciation of modern bear species was finished by the end of the Ice Age.
How long did this take? Traditionally, the creationist community followed Michael Oard’s model, which suggested from his mid-range calculations that the time until glacial maximum was about 500 years (with minimum and maximum values ranging from 174 to 1765 years) (Oard 2004). Further post-Flood climate modeling research by Dr. Steve Gollmer, however, starts with a cooler ocean model, and uses recent climate modeling technology. Gollmer’s research is ongoing, but appears to expose serious problems with Oard’s modeling (Gollmer 2018; Garner and Wood 2021). The main issue is that surface temperatures need to be much cooler to engender ice accumulation on land at an Ice Age scale. This suggests that the Ice Age may have started later than Oard’s ‘mid-range’ thesis indicates.
Another avenue of research suggests that use of the Septuagint may offer a more accurate post-Flood chronological timeframe. A number of papers have been published on this subject (Associates for Biblical Research 2019-2021), which essentially posits the Flood at about 3300 B.C., the Tower of Babel at about 2850 B.C., and God’s calling of Abraham at 2091 B.C. The Ice Age didn’t have as great an influence on the Middle Eastern landscape, so the Bible doesn’t specifically reference it. This timeframe does, however, give more time for the rapid adaptation, speciation, and migration events that occurred within the bear kind, within a creationist framework.
Evolutionists require long periods of time for the diversification of bears. Within a secular timetable, the Miocene period began about 23 million years ago. They require this great amount of time for the mutagenesis of beneficial traits and subsequent spread throughout populations. Creationists don’t require millions of years (or even hundreds of thousands of years), because in our model the created kind already has much of the necessary information for adaptation in its genome (termed ‘front-loading’ by some). Environmental cues may trigger the emergence of optional traits, which then may be selected for. Some creationists suggest that mutations aren’t always random; they may occur more frequently at genetic ‘hot spots’ which have a greater chance of producing beneficial developments.
Even secularists recognize that significant phenotypic changes don’t always reflect massive genetic change. Polar bears, for example, feed on blubber and flesh, and don’t need massive molars for grinding bones. Their reduced pre-molars and molars like developed rapidly through “the action of a few small mutations or simple developmental mechanisms” (Slater et al. 2010). In their examination of polar bear genetics, Castruita, Westbury, and Lorenzen (2020) found evidence for selective pressures on both standing variation within the brown bear ancestry and on de novo mutations in the polar bear genome. Biallelic sites in the polar bear’s ancestry provided alternative pathways for amino acid production that could influence such traits as lipid-binding proteins (allowing polar bears to adapt to a lipid-rich diet) and lack of fur pigmentation.
Introgression (transfer of genes through hybridization) was also important. Lan, et al. (2016) noted, “Our data suggest that following the divergence between ancestors of black, brown, and polar bears, introgression events among these species involved significant gene flow into the Arctic lineage at least twice from ancestors of extant brown bears and once from ancestors of extant black bears, possibly facilitating the capture of novel genes by Arctic specialists (polar bears) from colonizing boreal generalists (brown and black bears). Although there is likely strong purifying selective pressure on polar bear phenotypic features adapted to extreme Arctic life, novel, heritable traits transferred to polar bears could have become selectively advantageous during certain past periods of climatic change.”
There are several creation biologists who continue working on details involving rapid diversification within created kinds. I recommend a look at Jeanson (2017) and Carter (2021) for pertinent discussion.
THE BEAR KIND
A summary of diversification of bears after the Flood.
Amphicynodontinae and Hemicyoninae may or may not be
within the monobaramin. See those sections for discussion.
The eight living species are noted in red.
References
Abella, J., et al. 2012. Kretzoiarctos gen. nov., the oldest member of the giant panda clade. PLOS One 7(11): e48985.
Abella, J., et al. 2019. The last record of an ailuropod bear from the Iberian Peninsula. Geodiversitas 41(23): 797-809.
Arment, C. 2020a. To the Ark, and back again? Using the marsupial fossil record to investigate the post-Flood boundary. Answers Research Journal 13: 1-22.
Arment, C. 2020b. Implications of creation biology for a Neogene-Quaternary Flood/Post-Flood Boundary. Answers Research Journal 13: 241-256.
Asakura, S. 1969. A note on a bear hybrid Melursus ursinus x Helarctos malayanus at Tama Zoo, Tokyo. International Zoo Yearbook 9(1): 88.
Associates for Biblical Research. 2019-2021. The Genesis 5 and 11 Genealogy and Chronology Research Project. https://biblearchaeology.org/abr-projects/genesis-5-11-project
Barlow, A., et al. 2018. Partial genomic survival of cave bears in living brown bears. Nat. Ecol. Evol. 2(10): 1563-1570.
Baryshnikov, G. F., and A. V. Lavrov. 2013. Pliocene bear Ursus minimus Devèze de Chabriol et Bouillet, 1827 (Carnivora, Ursidae) in Russia and Kazakhstan. Russian Journal of Theriology 12(2): 107-118.
Baryshnikov, G. F., and P. A. Tleuberdina. 2017. Late Miocene Indarctos (Carnivora: Ursidae) from the Karabulak Formation of the Kalmakpai River (Zaisan Depression, Eastern Kazakhstan). Proceedings of the Zoological Institute RAS 321(1): 3-9.
Baryshnikov, G. F., and D. S. Zakharov. 2013. Early Pliocene bear Ursus thibetanus (Mammalia, Carnivora) from Priozernoe Locality in the Dniester Basin (Moldova Republic). Proceedings of the Zoological Institute RAS 317(1): 3-10.
Bocherens, H., et al. 2011. Niche partitioning between sympatric genetically distinct cave bears (Ursus spelaeus and Ursus ingressus) and brown bear (Ursus arctos) from Austria: Isotopic evidence from fossil bones. Quaternary International 245(2011): 238-248.
Bryant, H. N. 1993. Carnivora and Creodonta of the Calf Creek Local Fauna (Late Eocene, Chadronian), Cypress Hills Formation, Saskatchewan. Journal of Paleontology 67(6): 1032-1046.
Cahill, J. A., et al. 2013. Genomic evidence for island population conversion resolves conflicting theories of polar bear evolution. PLOS Genetics 9(3): e1003345.
Cahill, J. A., et al. 2015. Genomic evidence for geographically widespread effect of gene flow from polar bears into brown bears. Molecular Ecology 24: 1205-1217.
Calvignac, S., et al. 2008. Ancient DNA evidence for the loss of a highly divergent brown bear clade during historical times. Molecular Ecology 17: 1962-1970.
Carter, R. 2021. Species were designed to change, part 3: the tangled web of (intrabaraminic) life. CMI https://creation.com/species-designed-to-change-part-3
Castruita, J. A. S., M. V. Westbury, and E. D. Lorenzen. 2020. Analyses of key genes involved in Arctic adaptation in polar bears suggest selection on both standing variation and de novo mutations played an important role. BMC Genomics 21:543.
Czaplewski, N. J., and S. Willsey. 2013. Late Quaternary brown bear (Ursidae: Ursus cf. arctos) from a cave in the Huachuca Mountains, Arizona. In Gottfried, G., et al., eds. Merging Science Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III; 2012 May 1-5; Tucson, AZ. Proceedings. Fort Collins, CO: USDA, Forest Service, Rocky Mountain Research Station.
De Bonis, L. 2012. Ursidae (Mammalia, Carnivora) from the Late Oligocene of the ‘Phosphorites du Quercy’ (France) and a reappraisal of the genus Cephalogale Geoffroy, 1862. Geodiversitas 35(4): 787-814.
De Bonis, L., et al. 2017. A new late Miocene ailuropodine (giant panda) from Rudabánya (north-central Hungary). Geobios 50: 413-421.
De Torres Perezhidalgo, T. 1992. The European descendants of Ursus etruscus C. Cuvier (Mammalia, Carnivora, Ursidae). Boletin Geológico y Minero 103-4(632-642): 12-22.
Diffendal, R. F. 1990. The Sidney Gravel and Kimball Formation; Supposed parts of the Ogallala Group (Neogene), are not objectively mappable units. (University of Nebraska-Lincoln) Papers in Natural Resources 137: 23-38
Dong, W. 2008. Virtual cranial endocast of the oldest giant panda (Ailuropoda microta) reveals great similarity to that of its extant relative. Naturwissenschaften 95: 1079-1083.
Donohue, S. L., et al. 2013. Was the giant short-faced bear a hyper-scavenger? A new approach to the dietary study of ursids using dental microwear textures. PLOS One 8(10): e77531.
Everhart, M. J., and R. A. Hawkins. 2020. A giant bear mandible (Agriotherium schneideri Sellards 1916) from Sedgwick County, Kansas. Transactions of the Kansas Academy of Science 123(1-2): 191-202.
Figueirido, B., and L. H. Soibelzon. 2009. Inferring palaeoecology in extinct tremarctine bears (Carnivora, Ursidae) using geometric morphometrics. Lethaia 10.1111/j.1502-3931.2009.00184.x.
Figueirido, B., et al. 2011. Cranial shape transformation in the evolution of the giant panda (Ailuropoda melanoleuca). Naturwissenschaften 98: 107-116.
Fortes, G. G., et al. 2016. Ancient DNA reveals differences in behavior and sociality between brown bears and extinct cave bears. Molecular Ecology 25(19): 4907-4918.
Fowler, N. L., et al. 2021. Denning, metabolic suppression, and the realisation of ecological opportunities in Ursidae. Mammal Review https://doi.org/10.1111/mam.12246
Galbreath, G. J., et al. 2008. An apparent hybrid wild bear from Cambodia. Ursus 19(1): 85-86.
Gallo-Reynoso, J-P., et al. 2008. Probable occurrence of a brown bear (Ursus arctos) in Sonora, Mexico, in 1976. The Southwestern Naturalist 53(2): 256-260.
García, N., and J. L. Arsuaga. 2001. Ursus dolinensis: a new species of Early Pleistocene ursid from Trinchera Dolina, Atapuerca (Spain). Comptes Rendus de l'Académie des Sciences - Series IIA - Earth and Planetary Science 332(11): 717-725.
García-Rangel, S. 2012. Andean bear Tremarctos ornatus natural history and conservation. Mammal Review 42(2): 85-119.
Gardin, A., et al. 2021. Climbing adaptations of an enigmatic early Arctoid carnivoran: The functional anatomy of the forelimb of Amphicynodon leptorhynchus from the Lower Oligocene of the Quercy Phosphorites (France). Journal of Mammalian Evolution https://doi.org/10.1007/s10914-021-09553-w
Garner, P., and T. C. Wood. 2021. Ice Age (with Dr. Steve Gollmer). Let’s Talk Creation. https://www.youtube.com/watch?v=Evt9zZjid_g
Gollmer, S. M. 2018. Effect of aerosol distributions on precipitation patterns needed for a rapid Ice Age. In Proceedings of the Eighth International Conference on Creationism, ed. J.H. Whitmore, pp. 695-706. Pittsburgh, Pennsylvania: Creation Science Fellowship.
Gretzinger, J., et al. 2019. Large-scale mitogenomic analysis of the phylogeography of the Late Pleistocene cave bear. Scientific Reports 9: 10700.
Guilday, J. E. Grizzly bears from eastern North America. American Midland Naturalist 79(1): 247-250.
Hänni, C., et al. 1994. Tracking the origins of the cave bear (Ursus spelaeus) by mitochondrial DNA sequencing. Proceedings of the National Academy of Science USA 91(December): 12336-12340.
He, J-H., Z-B. Li, and Q. Wang. 2016. A new fractional derivative and its application to explanation of polar bear hairs. Journal of King Saud University 28: 190-192.
Hendey, Q. B. 1977. Fossil bear from South Africa. South African Journal of Science 73(April): 112-116.
Hennigan, T. 2010. The case for holobaraminic status in bears (Family Ursidae) and the implications within a creation model of ecology. CRSQ 46: 271-283.
Hofreiter, M., et al. 2002. Ancient DNA analyses reveal high mitochondrial DNA sequence diversity and parallel morphological evolution of Late Pleistocene cave bears. Mol. Biol. Evol. 19(8): 1244-1250.
Hunt, Jr., R. M. 1998. Ursidae. In: Janis, C.M. et al. Evolution of Tertiary Mammals of North America. Vol. 1. Cambridge, UK: Cambridge University Press.
Ingólfsson, Ó., and Ø. Wiig. 2008. Late Pleistocene fossil find in Svalbard: the oldest remains of a polar bear (Ursus maritimus Phipps, 1744) ever discovered. Polar Research 28: 455-462.
Jeanson, N. T. 2017. Replacing Darwin: The New Origin of Species. Green Forest, Arkansas: Master Books.
Jiangzuo, Q., and J. J. Flynn. 2020a. A new species of Agriotherium from North America, and implications for understanding transformations in the metaconid-entoconid complex of bears. Journal of Mammalian Evolution 27: 775-787.
Jiangzuo, Q., and J. J. Flynn. 2020b. The earliest ursine bear demonstrates the origin of plant-dominated omnivory in Carnivora. iScience 23, 101235.
Jiangzuo, Q., and R. C. Hulbert Jr. 2021. Coexistence of Indarctos and Amphimachairodus (Carnivora) in the Late Early Hemphillian of Florida, North America. Journal of Mammalian Evolution https://doi.org/10.1007/s10914-021-09546-9
Jin, C., et al. 2007. The first skull of the earliest giant panda. Proceedings of the National Academy of Science USA 104(26): 10932-10937.
Khattab, M. Q., and H. Tributsch. 2015. Fibre-optical light scattering technology in polar bear hair: A re-evaluation and new results. Journal of Advanced Biotechnology and Bioengineering 3(2): 1-14.
Knapp, M., et al. 2009. First DNA sequences from Asian cave bear fossils reveal deep divergences and complex phylogeographic patterns. Molecular Ecology 18: 1225-1238.
Knapp, M. 2019. From a molecules’ perspective—contributions of ancient DNA research to understanding cave bear biology. Historical Biology 31(4): 442-447.
Koon, D. W. 1998. Is polar bear hair fiber optic? Applied Optics 37(15): 3198-3200.
Krause, J., et al. 2008. Mitochondrial genomes reveal an explosive radiation of extinct and extant bears near the Miocene-Pliocene boundary. BMC Evolutionary Biology 8:220.
Kumar, V., et al. 2017. The evolutionary history of bears is characterized by gene flow across species. Scientific Reports 7: 46487.
Kutschera, V. E., et al. 2004. Bears in a forest of gene trees: Phylogenetic inference is complicated by incomplete lineage sorting and gene flow. Mol. Biol. Evol. 31(8): 2004-2017.
Lammers, F., et al. 2017. Phylogenetic conflict in bears identified by automated discovery of transposable element insertions in low-coverage genomes. Genome Biol. Evol. 9(1): 2862-2878.
Lan, T., et al. 2016. Genome-wide evidence for a hybrid origin of modern polar bears. Preprint. bioRxiv.org https://www.biorxiv.org/content/10.1101/047498v1
Laurie, A., and J. Seidensticker. 1977. Behavioral ecology of the sloth bear (Melursus ursinus). Journal of Zoology, London 182: 187-204.
Li, F., et al. 2017. Rediscovery of the sun bear (Helarctos malayanus) in Yingjiang County, Yunnan Province, China. Zoological Research 38(4): 206-207.
Lindqvist, C., et al. 2010. Complete mitochondrial genome of a Pleistocene jawbone unveils the origin of polar bear. Proceedings of the National Academy of Science USA 107(11): 5053-5057.
Liu, S., et al. 2014. Population genomics reveal recent speciation and rapid evolutionary adaptation in polar bears. Cell 157(May 8): 785-794.
Martin, J. E. 2013. A late occurrence of the bear Agriotherium from the Blancan Ringold Formation in southeastern Washington. Proceedings of the South Dakota Academy of Science 92(2013): 123-126.
Mayr, E. 1986. Uncertainty in science: Is the giant panda a bear or a raccoon? Nature 323: 769-771.
McLellan, B., and D. C. Reiner. 1994. A review of bear evolution. Int. Conf. Bear Res. And Manage. 9(1): 85-96.
McNab, B. K. 1992. Rate of metabolism in the termite-eating sloth bear (Ursus ursinus). Journal of Mammalogy 73(1): 168-172.
Meijaard, E. 2004. Craniometric differences among Malayan sun bears (Ursus malayanus); evolutionary and taxonomic implications. The Raffles Bulletin of Zoology 52(2): 665-672.
Miller, W. E., and O. Carranza-Castañeda. 1996. Agriotherium schneideri from the Hemphillian of Central Mexico. Journal of Mammalogy 77(2): 568-577.
Mitchell, K. J., et al. 2016. Ancient mitochondrial DNA reveals convergent evolution of giant short-faced bears (Tremarctinae) in North and South America. Biology Letters 12: 20160062.
Mondolfi, E., and E. O. Boede. 1981. A hybrid of a spectacled bear (Tremarctos ornatus) and an Asiatic black bear (Selenarctos thibetanus) born at the Maracay Zoological Park, Venezuela (Mammalia; Ursidae). Memoria de la Sociedad de Ciencias Naturales La Salle 41: 143-148.
Morlo, M., and D. Nagel. 2007. 7. The carnivore guild of the Taatsiin Gol area: Hyaenodontidae (Creodonta), Carnivora, and Didymoconida from the Oligocene of Central Mongolia. In: Daxner-Höck, G. (ed.) Oligocene-Miocene Vertebrates from the Valley of Lakes (Central Mongolia): Morphology, Phylogenetic and Stratigraphic Implications. Ann. Naturhist. Mus. Wien 108 A: 217-231.
Nakagome, S., S. Mano, and M. Hasegawa. 2013. Ancestral polymorphisms and sex-biased migration shaped the demographic history of brown bears and polar bears. PLOS One 8(11): e78813.
Oard, M. 2004. The peak of the Ice Age. AiG https://answersingenesis.org/environmental-science/ice-age/peak-of-ice-age/
O’Brien, S. J., et al. 1985. A molecular solution to the riddle of the giant panda’s phylogeny. Nature 317: 140-144.
Ogino, S., et al. 2011. New species of Agriotherium (Mammalia, Carnivora) from the late Miocene to early Pliocene of central Myanmar. Journal of Asian Earth Sciences 42(3): 408-414.
Pongracz, J. D., et al. 2017. Recent hybridization between a polar bear and grizzly bears in the Canadian Arctic. Arctic 70(2): 151-160.
Prasad, K. N. 1996. Pleistocene cave fauna from peninsular India. Journal of Caves and Karst Studies 58(1): 30-34.
Preuß, A., et al. 2009. Bear-hybrids: behavior and phenotype. Der Zoologische Garten N.F. 78: 204-220.
Puckett, E. E., et al. 2015. Phylogeographic analyses of American black bears (Ursus americanus) suggest four glacial refugia and complex patterns of postglacial admixture. Molecular Biology and Evolution 32(9): 2338-2350.
Qiu, Z. 2003. Chapter 2: Dispersals of Neogene carnivorans between Asia and North America. Bulletin of the American Museum of Natural History 2003(279): 18-31.
Qiu, Z., T. Deng, and B. Wang. 2009. First ursine bear material from Dongxiang, Gansu. Vertebrata PalAsiatica 47(4): 245-264.
Qiu, Z., T. Deng, and B. Wang. 2014. A Late Miocene Ursavus skull from Guanghe, Gansu, China. Vertebrata PalAsiatica 52: 265-302.
Rinker, D. C., et al. 2019. Polar bear evolution is marked by rapid changes in gene copy number in response to dietary shift. PNAS 116(27): 13446-13451.
Salesa, M. J., et al. 2006a. Anatomy of the “false thumb” of Tremarctos ornatus (Carnivora, Ursidae, Tremarctinae): phylogenetic and functional implications. Estudios Geológicos 62(1): 389-394.
Salesa, M. J., et al. 2006b. Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas. Proceedings of the National Academy of Science USA 103(2): 379-382.
Salis, A. T., et al. 2021. Ancient genomes reveal hybridisation between extinct short-faced bears and the extant spectacled bear (Tremarctos ornatus). Preprint. bioRxiv https://www.biorxiv.org/content/10.1101/2021.02.05.429853v1.full
Samuels, J. X., et al. 2009. The first mid-Blancan occurrence of Agriotherium (Ursidae) in North America: A record from Hagerman Fossil Beds National Monument, Idaho. Journal of Paleontology 83(4): 597-603.
Scherren, H. 1907. Some notes on hybrid bears. Proceedings of the Zoological Society of London 77(2): 431-435.
Schubert, B. W., et al. 2010. Giant short-faced bears (Arctodus simus) in Pleistocene Florida USA, a substantial range extension. Journal of Paleontology 84(1): 79-87.
Schubert, B. W., et al. 2019. Yucatán carnivorans shed light on the Great American Biotic Interchange. Biology Letters 15: 20190148.
Schultz, C. B., and L. D. Martin. 1975. Part 4. Bears (Ursidae) from the Late Cenozoic of Nebraska. Bulletin of the University of Nebraska State Museum 10(1): 47-54.
Seidensticker, J., K. Yoganand, and A. J. T. Johnsingh. 2011. Sloth bears living in seasonally dry tropical and moist broadleaf forests and their conservation. In: McShea, W. J., et al. eds., The Ecology and Conservation of Seasonally Dry Forests in Asia. Washington, DC: Smithsonian Institution Scholarly Press, pp.217-236.
Shao, Z., Y. Wang, and H. Bai. 2020. A superhydrophobic textile inspired by polar bear hair for both in air and underwater thermal insulation. Chemical Engineering Journal 397: 125441.
Shields, G. F., et al. 2000. Phylogeography of mitochondrial DNA variation in brown bears and polar bears. Molecular Phylogenetics and Evolution 15(2): 319-326.
Slater, G. J., et al. 2010. Biomechanical consequences of rapid evolution in the polar bear lineage. PLOS One 5(11): e13870.
Soibelzon, L. H., and B. W. Schubert. 2011. The largest known bear, Arctotherium angustidens, from the Early Pleistocene Pampean Region of Argentina: With a discussion of size and diet trends in bears. Journal of Paleontology 85(1): 69-75.
Soibelzon, L. H., E. P. Tonni, and M. Bond. 2005. The fossil record of South American short-faced bears (Ursidae, Tremarctinae). Journal of South American Earth Sciences 20: 105-113.
Soibelzon, L. H., et al. 2008. A Blancan (Pliocene) short-faced bear from El Salvador and its implications for Tremarctines in South America. N. Jb. Geol. Paläont. Abh. 250/1: 1-8.
Sorkin, B. 2006. Ecomorphology of the giant short-faced bears Agriotherium and Arctodus. Historical Biology 18(1): 1-20.
Sotnikova, M., et al. 2021. New species of Ballusia Ginsburg and Morales, 1998 (Ursidae, Carnivora) from Miocene of Eastern Siberia, Russia. Historical Biology 33(4): 486-497.
Stiller, M., et al. 2014. Mitochondrial DNA diversity and evolution of the Pleistocene cave bear complex. Quaternary International 339-340(August 7): 224-231.
Stucchi, M., et al. 2009. A 6,000+ year old specimen of a spectacled bear from an Andean cave in Peru. Ursus 20(1): 63-68.
Stynder, D. D., et al. 2019. A dental microwear texture analysis of the Early Pliocene African ursid Agriotherium africanum (Mammalia, Carnivora, Ursidae). Journal of Mammalian Evolution DOI: 10.1007/s10914-018-9436-y
Tedford, R. H., and J. Martin. 2001. Plionarctos, a tremarctine bear (Ursidae: Carnivora) from western North America. Journal of Vertebrate Paleontology 21(2): 311-321.
Terlato, G., et al. 2019. Chronological and isotopic data support a revision for the timing of cave bear extinction in Mediterranean Europe. Historical Biology 31(4): 474-484.
Trevino, J. C., and C. Jonkel. 1986. Do grizzly bears still live in Mexico? Bears: Their Biology and Management 6: 11-13.
Tseng, Z. J., C. Grohé, and J. J. Flynn. 2016. A unique feeding strategy of the extinct marine mammal Kolponomos: convergence on sabretooths and sea otters. Proceedings of the Royal Society B 283: 20160044.
Vallittu, P. K., et al. 2021. Temporomandibular joint and giant panda’s (Ailuropoda melanoleuca) adaptation to bamboo diet. Scientific Reports 11: 14252.
Van Gelder, R. G. 1977. Mammalian hybrids and generic limits. American Museum Novitates 2635: 1-25.
Van Valkenburgh, B. 1994. Extinction and replacement among predatory mammals in the North American Late Eocene and Oligocene: Tracking a paleoguild over twelve million years. Historical Biology 8: 129-150.
Viranta, S. 2004. Habitat preferences of European Middle Miocene omnivorous ursids. Acta Palaeontologica Polonica 49(2): 325-327.
Waits, L. P., et al. 1998. Mitochondrial DNA phylogeography of the North American brown bear and implications for conservation. Conservation Biology 12(2): 408-417.
Wallace, S. C., and X. Wang. 2004. Two new carnivores from an unusual late Tertiary forest biota in eastern North America. Nature 431 (September 30): 556-559.
Wang, X., M. C. McKenna, and D. Dashzeveg. 2005. Amphicticeps and Amphicynodon (Arctoidea, Carnivora) from Hsanda Gol Formation, Central Mongolia and phylogeny of basal arctoids with comments on zoogeography. American Museum Novitates (3843): 1-57.
Wang, X., et al. 2017. A basal ursine bear (Protarctos abstrusus) from the Pliocene High Arctic reveals Eurasian affinities and a diet rich in fermentable sugars. Scientific Reports 7: 17722.
White, T. 1965. The possibility of grizzly bears still existing in Saskatchewan. Blue Jay 23(3): 136-140.
Wood, T. C. 2021. Baraminology by cluster analysis: a response to Reeves. Appendix. Answers Research Journal 14 (https://assets.answersingenesis.org/doc/articles/arj/v14/reeves-response-appendix.pdf)
Wozencraft, W. C. 1989. The phylogeny of the recent carnivora. Ch. 18. In: Gittleman, J. L. (ed.), Carnivore Behavior, Ecology, and Evolution. Dordrecht: Springer Science.




